人間の記憶の仕組みをLLMに載せたい
LangChainのMemoryにはいくつかのタイプがある。
- 一定数のやり取りをバッファする
- 過去のやり取りを要約する
-
特定のEntityに関する情報を蓄積して利用する
- まだよく理解できていない
- Knowledge Graphで保存して利用する
- VectorStoreに保存して利用する
ただ、人間の記憶の仕組みは短期記憶、長期記憶などもっと複雑なはず。
その原理をちゃんと理解して、モデル化してLLMに搭載できるようにしたいのがゴール。
記憶の種類別にVectorStoreを用意しているプロジェクト。

記銘 → 保持 → 想起、というステップを踏むという理解らしい。
短期記憶は海馬、それ以外は大脳と大脳皮質?
目や耳などの感覚器から入ってきた情報は、脳の奥深くにある「海馬」という部位で一時的に保管されますが、そのほとんどは消え去ります(忘却)。
しかし、くり返し思い浮かべたり口に出したりすることによってかたちづくられた情報だけが、大脳皮質へ送られ、そこに刻みこまれるのです。
記憶は、保たれる時間によって「短期記憶」と「長期記憶」に分けられます。海馬の容量は小さいのですが、大脳皮質は大容量。海馬にとどまるだけの情報は、数秒から1分ほどの短い記憶(短期記憶)で、大脳皮質に送られた情報は、数分から年単位におよぶ長期記憶となります。
記憶は眠っているあいだに転送されるようで、そのプロセスが夢となってあらわれるのではないかとも考えられています。
海馬が短期記憶のBufferの役割をしていて、睡眠中に大脳に転送されて長期記憶化するらしい。
会話の文脈みたいな中期記憶の仕組みはないんだろうか?
記憶される情報には時間制限があり、繰り返し想起された記憶は長期記憶化される。
長期記憶の更新はリアルタイムじゃなくても良さそう。
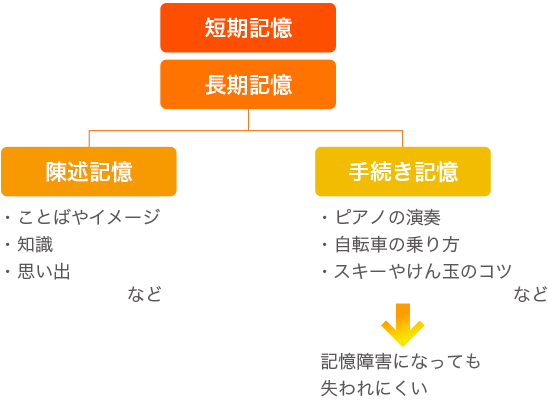
記憶は「短期記憶」と「長期記憶」に分けられます。さらに、長期記憶は、陳述記憶と手続き記憶に分けられます。
海馬をつかう陳述記憶は、ことばやイメージであらわすことのできるもので、学習による意味記憶(知識に関するもの)と体験によるエピソード記憶(主観的な思い出)があります。
手続き記憶というのは、たとえばピアノの演奏や自転車の乗り方、スキーの技術、けん玉のコツなど、からだで覚えた「動作や技能の記憶」。大脳基底核と小脳をつかうため、記憶障害になっても失われにくいと考えられます。
長期記憶には種類があって、種類が違うと使用する脳の部位も異なるらしい。
つまり異なるハード(VectorStoreなど)を使用するということか。
脳の部位と役割の違いでモデル化ができそう。
もう少し長期記憶の仕組みや中期的な記憶の仕組みがないのか調べたい。
海馬の詳しーい説明がされていそう。
海馬の英称は「Hippocampus」。
海馬は「海馬体(hippocampal formation)」とよばれる大脳辺縁系の一部である。海馬体は、歯状回(dentate gyrus)、海馬、海馬支脚(subiculum)、前海馬支脚(presubiculum)、傍海馬支脚(parasubiculum)、嗅内皮質(entorhinal cortex)に分けられる。このうち、歯状回、海馬、海馬支脚は、細胞層が単層であり、その上下を低細胞密度の層と無細胞層が挟んでいる。そのほかの部位は複数の層からなっている。とりわけ歯状回と海馬にみられる単純な層構造は、神経解剖学や電気生理学の研究進展に貢献してきている。
層構造になっている。
海馬が魅力的である二つ目の理由は、1950年代前半ごろから海馬がある種の記憶や学習に基本的な役割を演じていることが認知されるようになったからである。とりわけ、1957年に出されたScovilleとMilnerの報告は神経心理学に重要な一石を投じた。これはHMというイニシャルをもつ患者の報告である。おそらくHMは神経心理学の分野ではもっとも詳しく検査された人物である。彼はてんかんの治療の目的で両側の海馬を取り除く手術を受けたが、その後、新しい情報を長期記憶に留める能力が永遠に欠如してしまったのだ。この発見を機に、海馬は、記憶・学習の脳内メカニズムを理解しようという風潮から、神経解剖学、生理学、行動学などの分野で盛んに研究されるようになった。現在では海馬と記憶の関係は疑いのないものとなっている。
海馬が記憶に関連するのは解剖学的に確認されている。
海馬は、解剖学的および機能的なその構造から、ほかの大脳皮質システムのモデルにもなっている。大脳皮質は最近5年ほどはよく研究されるようになってきてはいるものの、現在知られている中枢神経系のシナプス伝達に関する知見の多くは海馬体の研究に基づいたものである。海馬体に関する過去の知見の大多数は、歯状回か海馬を標本としている。そこで以下では海馬(それもラットの海馬)と歯状回について焦点を当てる。
長期記憶の理解にも重要になりそう。
介在細胞(または内因細胞)は古来、局所に集中した軸索叢(plexus)を持ち、GABAを放出し、樹状突起にスパインがない神経細胞として定義されている。細胞標識法や記録法などが進歩し、介在細胞は従来考えられていたよりもはるかに多様であることがわかり、伝統的な定義だけでは、どれも必ず例外が現れてしまう(Buckmaster and Soltesz, 1996)。ただ実際のところ、歯状回や海馬の介在細胞のほとんどは、シナプス標的を局所に持ち、スパインを欠き、GABA性である(Freund and Buzsaki, 1996)といって間違いない(図4)。
スパイン?
-> シナプスを繋いで神経回路の取りうる状態を増やす役割をするらしい。
GABA性?
-> GABAによる神経伝達のことらしい
海馬体のおおまかな神経回路はRamon y Cajal(1911)の時代から知られているが、その詳細は最近の研究によって解明されたものである(図5)。Andersenら(1971)は海馬体の各部位を一方向につなぐ特徴的な回路の重要性を主張し、これを「三シナプス性回路(trisynaptic circuit)」と名付けた。感覚情報の多くは嗅内皮質を通じて海馬に流れ込むので、嗅内皮質を三シナプス性回路のスタート地点と考えることが多い。嗅内皮質は、隣り合った二つの皮質領域から情報を受信している。すなわち、嗅周囲皮質(perirhinal cortex)と嗅後部皮質(postrhinal cortex)(霊長類では海馬傍回(parahippocampal cortex)と呼ばれる)が、複次的な感覚情報を嗅内皮質にリレーしている(Burwell, 2000)。この入力の多くは興奮性である(Martina et al., 2001)。膨大部後方皮質(retrosplenial cortex)もまた感覚情報の源となっているらしい(van Groen and Wyss, 1992, Wyss and van Groen, 1992)。
読めないけど回路構造も分かっているよう。

CA3再帰回路はまた「鋭波(sharp wave)」を生み出す部位でもある。これは少数のCA3錐体細胞グループが同期したバースト発火を起こすことによって惹起されると考えられている。この鋭波は、覚醒しながら安静している時、もしくは「徐波睡眠(slow-wave sleep)」の時に観察され、記憶形成に関係していると見られる(Buzsaki et al., 1989)。実際、再帰回路は連合記憶を可能にする基本構造であるといわれている(Kohonen, 1978)。この「パターンコンプリーション仮説」は、CA3錐体細胞のみでNMDA受容体遺伝子を除去したマウスによって、ごく最近、直接証明された。このマウスは、仮説から予測されるように、きわめて特定の記憶(つまり連合記憶)だけが障害を受けていた(Nakazawa et al., 2002)。
連合記憶?
-> 手続き記憶の別名らしい
7.海馬のシナプス可塑性
シナプス可塑性の一般的な定義は、シナプスの以前の活動に応じて、シナプス応答の大きさが変動することである。海馬の興奮性シナプスの多くは、様々なタイプの活動依存的なシナプス可塑性を示す。可塑性は、持続時間や時間経過などで、細かく分類される。以下、各々を見ていこう。
この章が記憶の振る舞いに関連しそう。
7-1 短期可塑性
短期可塑性は、持続時間が数百msから数分のものを指し、促通(facilitation)、テタヌス後増強(post-tetanic potentiation)、抑圧(depression)がある。
7-2 短期可塑性-促通
PPFの大きさは刺激の間隔によって変わる。50 msくらいの間隔の場合、EPSPは数百%にも増大されることがあるが、間隔が伸びるとPPFは減少していく。この減少はほぼ指数関数的減衰であり、時定数は100-200 ms程度である。
7-3 短期可塑性-テタヌス後増強
PTPには2つの成分があり。一つは5-10 secの減衰定数を持つ「増幅(augmentation)」と呼ばれる相であり、もう一つはもっと遅い減衰を示す、いわゆるPTPと呼ばれる相である(Johnson and Wu, 1995)。
7-4 短期可塑性-抑圧
こうした抑圧の時間経過は、放出される神経伝達物質の量に応じて、数百msから数分と様々である。もっと短時間(10 ms以下)で生じる抑圧もある。
短期記憶の時間的なスケールの参考になりそう。
7-5 長期可塑性
海馬のグルタミン酸作動性の興奮性シナプスには30分から数時間にわたって持続する長時間タイプのものも存在する。これは、シナプスを連発刺激するか、もしくはシナプス前細胞と後細胞を特定の時間パターンで刺激することで誘導される。まとめて、長期増強(LTP、long-term potentiation)や長期抑圧(LTD、long-term depression)などと呼ばれるが、そこには、いくつかの異なった分子メカニズムが含まれている。
長期記憶に関係しそう?
-> 「長期」の時間スケールが「30分から数時間」らしいので、むしろ中期的な記憶の仕組みっぽい。気になる。
7-6 長期可塑性-長期増強
誘導相のあとの時間帯を「発現相(expression phase)」と呼び、ここではテスト刺激への応答は、高頻度刺激前より50-200%ほど増強している。発現相が誘導相よりも長いことがLTPの特徴である。通常、誘導相は数秒から1分程度あるが、発現相は数日に及びうる。発現相がどこまで持続しうるかを調べるのは難しいが、LTPが永遠に続くということはないだろう。
通常、誘導相は数秒から1分程度あるが、発現相は数日に及びうる。
結構長い。
とりわけ、シナプス前線維とシナプス後細胞の両者を同時にシータ周期で連回刺激する方法(pairing)は、theta burst pairingと呼ばれ、さらに効率よくLTPを誘導できる。シータ周期の神経活動は実際に行動中の動物脳で観察されるので、こうしたパターン刺激は単なる高頻度刺激よりも生理的だと考えられている。
周期的な刺激が長期化しやすい?
LTPの重要な性質として、入力のあったシナプスに特異的に発生するというものがある(Hoffman et al., 2002)。言い換えれば、シナプス応答の変化が起こるのは、高頻度刺激やシータ刺激を受けたシナプスだけということである。またLTPには連合性(associative property)という特徴がある。これは、そのシナプスと同時に他のシナプスも活動したら、LTPが生じるという現象である。LTPはこうした特徴を併せ持つからこそ、記憶機構の候補として研究者の興味を惹きつけているのである(Bliss and Lynch, 1988; Madison et al., 1991; Teyler et al., 1994; Brown et al., 2002)。
同時に活動したシナプスがあると長期化しやすい現象か、確かに興味深い。
7-7 長期可塑性-長期抑圧
LTDの発現相も30分から数時間に渡って継続する。似たような現象として、LTPを起こしたシナプスに低頻度刺激を与えると、増強分が元に戻る「脱増強(depotentiation)」という現象も知られているが、これはおそらくLTDとは異なる機構によって生じるものである。多くの数理研究のモデルでは、特定のシナプス経路を増強させるために、他のシナプスを弱めることで学習を成立させている。つまり、LTD様の現象はずいぶんと昔から論理的には使われてきていたわけである。
抑圧を利用するという発想はなかった、なるほど。とはいえちゃんと理解できていないが。
こうしたデータから、外の世界を認識する地図(cognitive map)が海馬の中に形成されているものと推測されている(O’Keefe, 1979)。より一般的な意味では、海馬体の神経細胞は、様々に活性化されるユニットの組み合わせ、つまり「アセンブリー(assembly)」として働くことで、現在の経験を内部表象している、と考えることもできる。おそらく、こうした海馬の内部表象と、大脳皮質にあるより詳細な経験情報が相互作用することによって、長期的な記憶が形成されるのだろう(Wilson and McNaughton, 1993, 1994; McHugh et al., 1996)。これらの電気生理学的なデータが示唆することは、海馬体の神経細胞がある特定の情報に選択的に反応するわけではなく、むしろ、行動のすべてを表す内象を一時的に記憶しておく、いわば、短期記憶バッファーとして働いていると考察される。この内部表象が後に再生されることで、ゆっくりと大脳皮質の長期的な記憶に置き換えられていくのだろう(Eichenbaum, 2001; Haist et al., 2001)。実際、徐波睡眠(slow-wave sleep)中に海馬で、覚醒時での行動が内部再生されることはすでに示唆されている(Hoffmann and McNaughton, 2002)。
これが知りたかった考察!
より一般的な意味では、海馬体の神経細胞は、様々に活性化されるユニットの組み合わせ、つまり「アセンブリー(assembly)」として働くことで、現在の経験を内部表象している、と考えることもできる。
これらの電気生理学的なデータが示唆することは、海馬体の神経細胞がある特定の情報に選択的に反応するわけではなく、むしろ、行動のすべてを表す内象を一時的に記憶しておく、いわば、短期記憶バッファーとして働いていると考察される。
短期的な記憶もVectorのような表現(Embeddings)でBufferingしているのか、なるほど。
この情報を長期記憶に転送しているなら、当然長期記憶も同じ形式の情報を受け取って保持していることは想像できる。
短〜中期の記憶は海馬の働きでおおよそ理解できたので、次は長期記憶。
長期記憶の分類。
Credit:
鈴木 麻希
京都産業大学 コンピュータ理工学部 インテリジェントシステム学科
藤井 俊勝
東北福祉大学 感性福祉研究所 & 健康科学部
- 陳述記憶
- エピソード記憶
- 意味記憶
- 非陳述記憶
- 手続き記憶
- プライミング(以前の経験により、後に経験する対象の同定を促進(あるいは抑制)される現象)
- 古典的条件付け(パブロフの犬的なもの?)
- 非連合学習
分類が分かりやすくまとまっている。
もうちょっと脳の部位と記憶の種類の関連性が知りたい。
この論文の内容が気になる。
海馬は、エピソード記憶の形成や想起に重要な脳領域です。先行研究により、覚えた記憶は、時間経過とともに、海馬から大脳皮質に徐々に転送され、最終的には大脳皮質に貯蔵されるのではないかとのアイデアがありますが、大脳皮質への記憶の転送に関して、神経回路メカニズムの詳細はほとんど分かっていませんでした。
本研究では、記憶を担う細胞(記憶痕跡細胞またはエングラム細胞)を標識・操作する研究手法注1)を用いて、大脳皮質の前頭前皮質で、学習時に既にエングラム細胞が生成されていることを発見しました。この前頭前皮質のエングラム細胞は、海馬のエングラム細胞の入力を受けることによって、学習後徐々に構造的・生理的・機能的に成熟することも発見しました。逆に、海馬のエングラム細胞は、時間経過とともに活動休止、脱成熟することが分かりました。つまり、これまで考えられてきた海馬から大脳皮質への記憶の転送のアイデアは、前頭前皮質のエングラム細胞の成熟と海馬のエングラム細胞の脱成熟により、記憶想起に必要な神経回路が切り替わることで説明できるようになりました。
詳しく読みたい。
背景
海馬はエピソード記憶の形成や想起に重要な脳領域です。小動物を用いた海馬の損傷実験やヒトの症例研究によって、エピソード記憶の形成後、最初はその出来事を思い出すのに主に海馬を必要としますが、その記憶を覚えた後、時間経過に伴い徐々に海馬は必要でなくなり、数週間後には大脳皮質を使ってそのときの出来事を思い出すことが分かっていました。このことから、心理学者や脳科学者らは、“記憶”は、時間経過とともに、海馬から大脳皮質に徐々に転送され、最終的には大脳皮質に貯蔵されるのではないかと考えました。この考えは、「記憶固定化の標準モデル」と呼ばれています。
また、海馬から大脳皮質へと記憶想起に必要な脳領域が変化するに伴い、詳細な情報を含む体験的なエピソード記憶から、知識としての意味記憶へと記憶の性質が変わるのでないかとの考え方も提案され、大脳皮質への記憶固定化・記憶転送の実体やその仕組みを明らかにすることは、神経科学の分野で大きな課題の一つとなりました。
しかし、この問題にアプローチする研究のほとんどは、いつ、どの脳部位が記憶の思い出しに“必要”なのかを調べる損傷実験が主だったため、どの脳部位に記憶が“貯蔵”されているのか?そして、本当に海馬から大脳皮質へ記憶の転送は起きているのか?といった問題に対し、記憶を担っている細胞を標識する方法論がなかったために、直接的に答えることができずにきました。
ちょうど気になっていた部分。
つまり、この実験により、これまで考えられてきた記憶固定化の標準モデルとは異なり、学習時に活性化した前頭前皮質には、学習して1日後には既にエピソード記憶情報を保持している、エングラム細胞ができていることが分かりました。
エングラム細胞?
-> 記憶による神経回路の可塑性をもたらす細胞、という理解。
さらに、学習時における、海馬―大脳嗅内皮質から前頭前皮質への神経刺激(環境情報を提供する)と、恐怖記憶に関わる扁桃体から前頭前皮質への神経刺激(ショック情報を提供する)の両方が、前頭前皮質のエングラム細胞の生成には必須であることが分かりました。
重要そうだが、後者の神経刺激のモデル化はできるのか...?
関連論文も気になる。
では、学習時に前頭前皮質で生成されたエングラム細胞は、実際の記憶の想起時には使われるのか?そして、それは記憶想起に必要不可欠なのか?学習後1日では、前頭前皮質のエングラム細胞は実際の記憶の想起時にはあまり活動しませんでした(図1、学習後1日)。また、この時点では想起に必要でもありませんでした。しかし、学習後2週間では、この前頭前皮質のエングラム細胞は、他の細胞に比べ強く活動しました(図1、学習後2週間)。さらに、この前頭前皮質のエングラム細胞では、時間の経過とともに、樹状突起の数が増大して、2週間後には、記憶の想起に必要不可欠になりました。
つまり、前頭前皮質のエングラム細胞は、最初から記憶情報は持っているけれどもすぐには想起に使えない状態にあります。これを、「サイレントなエングラム」と呼ぶことにしました。しかし、時間経過とともに、樹状突起を増加させ、前頭前野での神経細胞同士のつながりを強化し、その結果、実際の記憶想起に使われるばかりでなく、そのために不可欠になりました。これらの実験結果は、前頭前皮質のエングラム細胞は、時間経過とともに、「成熟する」ことを示します。つまり、これらの細胞は「サイレント」な状態から「アクティブ」な状態に移行するということです。
さらに、研究チームは、海馬のエングラム細胞の神経出力を遮断する実験を行い、その結果、前頭前皮質のエングラム細胞の成熟には、海馬のエングラム細胞からの神経刺激の入力が必須であることを証明しました。
前頭前皮質のエングラム細胞は、サイレント状態から2週間後にアクティブな状態に変化する。
そのためには海馬のエングラム細胞からの刺激が必要。
これまでの研究によって、学習後1日の記憶想起には海馬のエングラム細胞が重要な役割を果たしていることが示されています注4)。さらに、上記のように、この海馬のエングラム細胞は、前頭前皮質のエングラム細胞の成熟に必要不可欠であることを示しました。しかし、学習後の時間経過に伴い、海馬で形成された記憶エングラムの運命はどうなっていくのでしょうか?
学習時に海馬で形成されたエングラム細胞は、記憶情報を保持し、確かに学習後1日は実際の記憶の想起にも使われます(図2、学習後1日)。しかし、学習後約2週間が経過すると、海馬のエングラム細胞の記憶情報は依然保たれているものの、実際の記憶の想起には活動しませんでした(図2、学習後2週間)。つまり、海馬のエングラム細胞は、前頭前皮質のエングラム細胞とは逆に、「アクティブ」な状態から、時間とともに「サイレント」な状態に移行することになります。
このサイレントエングラムに特別な機能があるかどうかを知るにはさらなる実験が必要です。しかし、一つには、サイレント状態を経て、最終的には海馬のエングラム細胞は完全に記憶情報を失う可能性があります。もしくは、サイレント状態は引き続き維持され、より詳細な記憶を想起しないといけない事態に出くわした際、そういった特別なときにのみ、再度、使われるのかもしれません。
海馬のエングラム細胞は2週間経過するとアクティブな状態からサイレントな状態に移行する。
サイレントな状態のエングラム細胞には記憶自体はあるが、想起には利用されない=思い出せない?
扁桃体のエングラム細胞は海馬のエングラム細胞と同様に、学習の直後にアクティブな状態で形成され、恐怖記憶に基づく「すくみ反応」に貢献し、また前頭前皮質のエングラム細胞の形成に不可欠です。本研究によって、さらに、学習後経過する時間にかかわらず、アクティブな状態を維持することが分かりました。ただし、記憶の想起に必要な環境情報を含む刺激は、学習後1週間程度においては、海馬および大脳嗅内皮質を通じて配達されますが、その後は、前頭前野のエングラム細胞を通じて配達されます。つまり、学習後に経過する時間にかかわらず、同じ扁桃体エングラムが使われるのですが、想起のための刺激の配達ルートにシフトが起こるということです。
扁桃体のエングラムは時間に関係なく成熟する。
扁桃体は恐怖や不安などのネガティブな情動を司るから、トラウマみたいな記憶は時間が経っても忘れないということ?
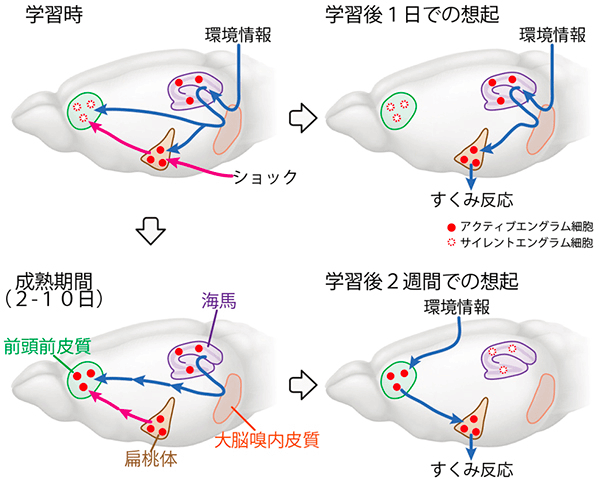
分かりやすい図があるのでこれを見ながら整理する。
- 学習時には、海馬と扁桃体ではエングラム細胞がアクティブな状態で生成される。この時、前頭前皮質ではエングラム細胞は生成されているがサイレントな状態で記憶の想起には使用されない。
- 学習後1日以内では、海馬→扁桃体と直接想起が行われ、前頭前皮質のエングラム細胞は使用されない。
- 学習後2~10日間では、海馬→前頭前皮質への神経入力が入ると、前頭前皮質のエングラム細胞がアクティブになり徐々に成熟する。
- 学習後2週間後、海馬のエングラム細胞はサイレント状態に戻り、前頭前皮質のエングラム細胞から直接想起がされる。
環境情報のエントリーポイントの判断はどこが行なっている?
想起の流れで整理するとこう?
- 海馬のエングラム細胞がない → 海馬のエングラム細胞をアクティブな状態で新規作成(学習開始) → 前頭前皮質のエングラム細胞をサイレントな状態で新規作成
- 海馬のエングラム情報ある&アクティブ&1日以内 → 海馬で想起する
- 海馬のエングラム情報ある&アクティブ&1日以降 → 海馬から前頭前皮質のエングラム細胞を成熟させつつ想起
- 前頭前皮質のエングラム細胞がアクティブ → 前頭前皮質で想起する
これならアルゴリズム化できそうでは。
ただこれだと海馬が前頭前皮質と同量のエングラム細胞をもつ必要があるが、それだと海馬の容量が多すぎるのでは?
この話を踏まえて先ほどの海馬の説明を見返してみたい。
エングラム関連の資料探し
詳しく読んでみる。
海馬は「いつ、どこで、何が起こった」という文脈情報に基づいたエピソード記憶に必要不可欠ですが、どのようなメカニズムでエピソード記憶を担うか、明らかではありません。今回、研究チームは、テトロード記録法[3]と光遺伝学[4]、そして特殊な遺伝子組換えマウスc-Fos-tTAマウス[5]を組み合わせ、マウスが新しい文脈を経験した際に形成される記憶エングラムの神経活動を記録することに成功しました。これまで、記憶エングラムには動物が経験した場所の位置情報が保存されていると予想されていましたが、解析の結果、記憶エングラムが表現する位置情報は極めて不安定であり、その活動は文脈のアイデンティティ[6](文脈を構成する情報の組み合わせ)に素早く応答していることが明らかになりました。これは、海馬には動物の位置情報を保存する場所細胞[7]とは別に、文脈情報を保存する記憶エングラムが存在することを示し、海馬が記憶エングラムの活動を通してエピソード記憶のインデックス(索引)として機能することを示しています。
海馬の記憶エングラムは文脈情報を持っていて、エピソード記憶におけるインデックスとして機能している、という研究。
一方で、「記憶インデックス仮説」という別の仮説も存在します。この仮説では、エピソード記憶を構成する情報は海馬ではなく大脳皮質に保存されていて、海馬にはそれらを呼び起こすためのインデックス(索引)が記録されているとしています。文脈条件づけ[8]という行動実験とc-Fosなどの最初期遺伝子[9]を使った研究では、この記憶インデックス仮説と矛盾しない結果が得られています。
例えば、動物が新しい文脈を経験するとき、海馬には、空間内を移動しなくても活性化される神経細胞が存在します。このように、場所に限らず文脈に応答して活性化される海馬神経細胞群は「記憶エングラム」とも呼ばれ、人為的に活動を誘導・抑制することでその文脈についての記憶を呼び起こしたり、抑制したりできることが近年の研究で分かってきました。さらに、海馬の記憶エングラムの活動を抑制することで、記憶を想起しようとする際に大脳皮質の神経細胞が再活性化されなくなるという報告も記憶インデックス仮説と合致します。
海馬と大脳皮質内のエピソード記憶の関連性にまつわる話のよう。
この結果は、記憶エングラムがその活動量の変化を通して、文脈のアイデンティティ(文脈を構成する情報の組み合わせ)を表現していることを示しています。
以上の結果から、海馬には、認知地図仮説が空間記憶の素子として想定する場所細胞と、文脈のアイデンティティを表す記憶エングラムの2種類の記憶痕跡が別々に存在することが分かりました。さらに、「記憶エングラムが記憶インデックス仮説の提唱する記憶インデックスの実体であり、その活動が動物の脳内で表現される経験の情報と結びつくことでエピソードを定義する役割を担っている」という仮説を提唱しました。
最後のコメント部分が気になる。
次に、このタンパク質を脳の様々な部位に導入すると、光を使って記憶を消すことができるようになりました。面白いことに、学習の直後に記憶ができるシナプスや、その後の睡眠中に記憶ができるシナプス、次の日の睡眠中に記憶ができるシナプスが、脳の異なった部位にそれぞれ存在することがわかりました。本研究により、記憶を長期間保つ、睡眠の機能の新しい細胞モデルを提唱し、記憶・睡眠研究の前進に貢献しました。
さらに記憶が皮質に移る時間枠を知るために、前帯状皮質での LTP 時間枠を調べたところ、学習の翌日の
睡眠中に前帯状皮質で LTP が誘導されていることが分かりました。つまり、長期的に保存されるための記憶
は学習の翌日には既に皮質に移行し始めていることが分かりました。
しばらく他の作業で手をつけられていないが、考えていることのメモだけ。
- 短期記憶も生のテキストではなくEmbeddingsで保持しているのでは?
- 複雑な長期記憶の仕組みは大変そうなので、まずはシンプルに考えるところから
- 記憶方法ばかりに目が行きがちだけど、睡眠と忘却も重要なファクターなのでは?